Sequential Order in Precise Phase Timing
 While many might accept that synchrony is an important part of neural computation, several others would be hesitant to do so. Many of these naysayers are wary of a "strong" form of the synchrony hypothesis, sometimes referred to as "polychrony," which suggests that information can be encoded by precise temporal relationships among multiple cell assemblies. Neurons are too simple, and the brain is too noisy (or so the logic goes) for such precision to be manageable. I tacitly agreed with this perspective until reading two fascinating articles in today's issue of Neuron.
While many might accept that synchrony is an important part of neural computation, several others would be hesitant to do so. Many of these naysayers are wary of a "strong" form of the synchrony hypothesis, sometimes referred to as "polychrony," which suggests that information can be encoded by precise temporal relationships among multiple cell assemblies. Neurons are too simple, and the brain is too noisy (or so the logic goes) for such precision to be manageable. I tacitly agreed with this perspective until reading two fascinating articles in today's issue of Neuron.Dragoi & Buzsaki recorded from 256 different neurons in rat hippocampus as the rats wandered around a track in search of a food reward. They recorded from two regions in particular: the CA1, and the CA3 regions of hippocampus. These regions are important because they are critically involved in the pattern separation (CA3) and pattern completion (CA1) processes that allow for effective memory storage and retrieval, respectively.
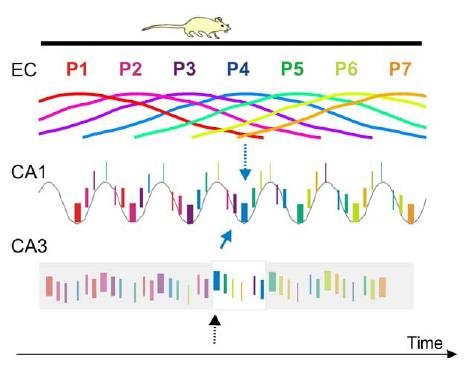
Before moving on, it's important to know that neurons in CA3 fire only in extremely specific physical locations. For example, a given CA3 neuron may respond to a specific range of movement, in a particular direction in a particular place, and for nothing else. CA1 neurons are slightly less specific, but still far moreso than neurons elsewhere in the hippocampal formation. The range of locations for which a CA3 or CA1 neuron will fire maximally is known as its "place field peak."
As shown previously by other research, the authors found that "the distance between adjacent place field peaks is represented by the precise temporal relations of spikes at a compressed or "theta" time scale on the order of milliseconds." In other words, the firing patterns of specific neurons would lag from the dominant local frequency (think of it as the overall "pulse" at this portion of the network) by an amount that is directly related to the physical distance between the rat and the place field peak for that neuron. Even more surprising is that by examining the phases of all pairs of the 256 neurons relative to the dominant hippocampal theta frequency, one can derive a kind of "temporal map" which relates the distance between each physical location in terms of phase lag in a single theta cycle.
Interestingly, the phases of CA3 & CA1 cell assemblies relative to the dominant theta frequency seems to relate to physical location such that previously visited locations are encoded by neurons on the descending side of the "theta trough," locations to which the rat appears to be heading are encoded by neurons on the ascending side of the theta trough, and in the very center of the theta phase are the neurons that encode the rat's current location. A process of phase precession serves to coax the firing rhythms from the ascending side into the trough of the theta rhythm, as the rat moves through its environment.
This research is important because it clarifies 1) how theta-clocked cycles of interaction between CA1 and CA3 regions might serve to consolidate temporal sequences of information into single "episodes"; 2) the mechanism for storage and retrieval of sequential order information that may underlie both spatial navigation and episodic memory, the two tasks most strongly associated with the hippocampus; 3) the mechanism that might be capable of generating the specific spike train patterns known to drive spike timing dependent plasticity.
Related Posts:
Polychrony
posted by Chris Chatham at 8:04:00 PM. Now go to the current Developing Intelligence blog!
![]()
![]()


0 Comments:
Post a Comment
<< Home